|
|
Subjekte Menschen können nur als Menschen sein, indem sie einander Subjekte sind. |
|
| Paradigmata |
• |
|
|
|
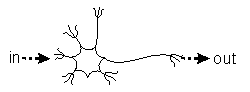
Der neurophysiologische AspektDas aktuelle Paradigma der Neurophysiologie wird am
klarsten, wenn man das Modell der Nervenzelle als ihrem grundlegenden
Untersuchungsgegenstand betrachtet (Abbildung 1). In diesem Paradigma kann
die Nervenzelle nur reagieren, autonome Aktionen der Nervenzellen sind in
diesem Konzept nicht denkbar. Auch beliebig viele zusammen geschaltete
Nerverzellen dieser Art können kein autonomes System ergeben. "Die völlige
Unbegründetheit dieser Anschauungen konnte deshalb nie zutage treten, weil
kein Reflexologe und kein Behaviorist je in die Lage kam, den Ablauf einer
längeren, hochdifferenzierten Kette arteigener angeborener
Verhaltensweisen überhaupt zu Gesicht zu bekommen. Die Forschungsmethode
beider großen mechanistischen Schulen beschränkte sich bekanntlich auf
Experimente, in denen eine Zustands-Änderung in den auf den
Organismus einwirkenden Umgebungsbedingungen gesetzt und die Antwort
des Tieres auf diese Änderung registriert wurde. Die vorgefaßte Meinung,
daß der Reflex und der bedingte Reflex die einzigen wesentlichen
„Elemente“ alles tierischen und menschlichen Verhaltens seien, bestimmte
eben eine ganz spezielle, kaum je variierte Art der Versuchsanordnung, bei
der das untersuchte Zentralnervensystem gewissermaßen gar keine
Gelegenheit bekam, zu zeigen, daß es auch etwas anderes zu leisten
imstande sei, als einwirkende Außenreize zu beantworten. Bei der
ausschließlichen Anwendung dieser Methodik musste die Meinung
bestärkt werden, daß sich die Leistung des Zentralnervensystems im
Aufnehmen und Beantworten äußerer Reize erschöpfe. In diese Richtung weisen auch die von von Holstund Mittelstae und Anochin vorgebrachten
kritischen Einwände gegen die Reflextheorie des Verhaltens. Sie
untersuchten, ob es unter den neuronalen Prozessen auch solche gibt, die
nicht dem Schema des Reflexes folgen und auch nicht mit diesem vereinbar
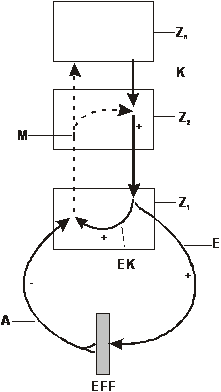
sind. „Schließlich dürfte der rhythmische Kontraktionsimpuls in einem Stück Bauchmark, das man aus dem Wurm herauspräpariert und in physiologische Lösung legt, nicht mehr zustande kommen, da hier ja alle Möglichkeiten eines Reflexreizes genommen sind – in Wirklichkeit läuft in solch einem isolierten Bauchmarkstück der Rhythmus pausenlos ab. Durch Ableitung der elektrischen Entladungen der Ganglienzellen lässt sich das Weiterlaufen des Rhythmus noch über Stunden nachweisen (unpubliziert). Der zentrale Rhythmus bedarf also überhaupt nicht notwendig peripherer Reize, er ist nicht reflektorischer, sondern automatischer Natur.“ (, S. 8f.) Zusammenfassend führt er aus: „Die zentrale Koordination beruht nicht auf Kettenreflexmechanismen, sie ist von grundsätzlich anderer Art. Ihre Werkzeuge sind Vorgänge, die sich allein im Innern des ZNS selbst abspielen. Der „Reflex“ ist dazu da, diesen inneren Ablauf den sich ändernden peripheren Bedingungen jeweils anzupassen, ihn nach dieser oder jener Richtung hin abzuwandeln. Er ist nicht der Grundvorgang selbst, wie man so allgemein glaubt, sondern entweder ein zusätzliches Attribut des zentralen Mechanismus oder, wohl meistens, ein kompliziertes Zusammenspiel zusätzlicher Mechanismen mit den hier beschriebenen aktiven zentralen Kräften.“ (Ebenda, S. 37) Von Holst fasst seine Kritik am traditionellen Reflexbegriff mit folgenden Worten zusammen: „Das kennzeichnende Merkmal dieser neuen Denkart ist eine Blickwendung um 180°. Wir fragen nicht nach der Beziehung zwischen einer gegebenen Afferenz und der durch sie bewirkten Efferenz, also nach dem „Reflex“, sondern gehen umgekehrt von der Efferenz aus und fragen: Was geschieht im ZNS mit der von dieser Efferenz über die Effektoren und Rezeptoren verursachten Afferenz, die wir die „Reafferenz“ nennen wollen?“ (Ebenda, S. 39) Die in Abbildung 2 wieder gegebene Darstellung
des Reafferenzprinzips von Holsts erfordert zumindest in Zn
eine Nervenzelle, die die autonom Impulse generiert (Abbildung 1 On
Mouseover). „Somit besitzen alle funktionellen Systeme
des Organismus ein und dieselbe prinzipielle physiologische Architektur.
Ihr Unterschied besteht nur in der Bestimmungstechnik. Ein System mit
einem stabilen lebenswichtigen Endeffekt (wie z. B. der osmotische Druck
im Blut) benutzt als Vergleichsapparat die erblich vorgegebenen
Eigenschaften entsprechender lebender Zellen, wodurch auch die Art der
Auslese der Zwischenaktionen bestimmt wird. Bei den episodisch
auftretenden funktionellen Systemen vom Verhaltenstyp hingegen bildet sich
der Vergleichsapparat (der Aktionsakzeptor) zusätzlich jedes Mal neu auf
der Grundlage der Afferenzsynthese aller im betreffenden Augenblick
vorhandenen inneren und äußeren Einwirkungen auf den Organismus. Sowohl von Holst als auch Anochin bleiben mit ihrer Terminologie im Rahmen des Behaviorismus und gestalten die Ergebnisse ihrer Arbeiten so als kumulativen Beitrag zu diesem Paradigma. Deshalb vermögen auch die Reafferenzprinzipien nicht das Korsett des Reflexkonzeptes zu verlassen. Die Formulierung eines neuen Paradigmas erfordert also auch eine eigenständige Terminologie. Um das hier entwickelte Begriffssystem nicht nachträglich wieder in dieses Korsett zu zwingen, werde ich in meiner Darstellung wie Ausdrücke „Reiz“ und „Reaktion“ nicht zur Beschreibung tierischen oder menschlichen Verhaltens benutzen. Der Behaviorismus führt sowohl in der Verhaltensbiologie wie in der Neurophysiologie zu einer Auffassung des Lebens, in der das Lebewesen auf das Niveau eines Automaten reduziert wird, der auf eine äußere Einwirkung, den Reiz, wartet, auf den er dann in bestimmter angeborener oder erlernter Weise reagiert. In dieser Sichtweise agiert das Lebewesen nicht, es reagiert. |
|
|
|
Weiterführende Links: Nervenzelle, |
|
|
Weiterführende Literatur: Lorenz, Konrad (1992): Über tierisches und menschliches Verhalten - Gesammelte Abhandlungen I und II, Piper & Co.Verlag, München, Zürich, Holst, Erich von (1974): Zentralnervensystem , Deutscher Taschenbuch Verlag GmbH & Co. KG, München, Anochin, Pjtor Kusmitsch (1978): Beiträge zur allgemeinen Theorie des funktionellen Systems, Gustav Fischer Verlag, Jena, |
||